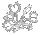

Nepenthes tenax, Queensland, Austraila. Photo © Richard Nunn.
The trouble with understanding the evolution of Nepenthes and its spectacular pitchers is in spite of the fact there are well in excesses of 100 species of Nepenthes today, the genus as a whole has no close relatives. And within the genus itself there are no obvious ancestral types or transitional species; no proverbial missing links. We are essentially presented with a finished product.
Nepenthes does have distant relatives that can give us hints about where the genus came from. But those relatives have had on the order of 60 million years to develop their own special adaptations since their split from the future Nepenthes. The carnivore Triphyophyllum and its non-carnivorous relatives Ancistrocladus, Habropetalum and Dioncophyllum, are jungle lianas strikingly like pitcherless Nepenthes when small. Unlike Nepenthes, the carnivorous leaves of Triphyophyllum are sundew-like. Drosophyllum, another distant relative to Nepenthes, has been confused with sundews. The fun part of this is if you remove most of the Nepenthes characters from Triphyophyllum you end up with Drosophyllum.
Even more distant relatives of Nepenthes are Drosera, the true sundews, and the snap-trap carnivores Dionaea and Aldrovanda. There are characters of these species that tie them to Nepenthes but not necessarily the carnivory aspect.
We know the way all of these genera are related based on DNA sequence data. For this phylogenetic analysis we are using the chloroplast trnK gene intron which contains the matK gene. If you do not know what that means, the important point is it is a different DNA sequence from the Drosera phylogeny. It is still a chloroplast sequence and not a nuclear sequence. So on the one hand we can compare results between different DNA sequences but on the other we only have matK sequences of a few Drosera species and we only have rbcL sequences of a few Nepenthes species. On the Drosera phylogeny we found Drosera arcturi linking into the phylogeny very close to the split of Drosera with its closest relatives. Drosera arcturi is a primitive Drosera by any measure and it has many attributes one could imagine the first sundews had. We have nothing like that for the Nepenthes phylogeny.

Nepenthes rafflesiana pitcher.
The data used for this tree are chloroplast trnK gene intron, matK sequences deposited with NCBI. Care was taken to exclude the translocated matK pseudogene sequences in the figure. Analysis was via NCBI Blast, graphed via PHYLIP drawtree unrooted tree drawing program and detailed manually in a drawing program. The lengths of the lines are an indication of genetic distance. Plumbago auriculata was used as the outgroup. Specific details of the phylogeny could change with additional species or analysis with a different program.
Nepenthes has a pompom-like phylogenetic tree. So do Triphyophyllum and its related genera as well as the genus Ancistrocladus. That is, we see a long arm of one species then a poof of many species bloom out. In the case of Nepenthes it is over a hundred species, Ancistrocladus has over 20 species, the Triphyophyllum clade has 3 very closely related monotypic genera. That does not mean only one Nepenthes or proto-Nepenthes species existed. It means of the possibly hundreds of potential proto-Nepenthes species, only one survived to produce what we know of today as Nepenthes. The same would go for the other 2 clades as well as Drosophyllum. This of course assumes there isn't some plant out there we have not discovered or tested related to Nepenthes. And since we are actually tracking chloroplast lineages what we really know is the history of the chloroplast lineage that survived. Via hybridization and polyploidy there could be many nuclear lineages combined and preserved.
The most basal species of the Nepenthes phylogeny is Nepenthes distillatoria. There is nothing primitive about this species relative to other Nepenthes species. And there is nothing primitive about N. pervillei, N. madagascariensis, N. masoalensis, and N. khasiana, the other basal species. All of these known species are separated from the actual ancestor by many genetic changes in matK and presumably other genes but in terms of what makes them a Nepenthes there is nothing to make one think they are "ancestral" just looking at them. We would presume then that if we were to find a fossil of the Nepenthes species that gave rise to basal species it would be fully modern.
These basal species plus the near-basal N. danseri, N. neoguineensis, N. tomoriana and N. paniculata plus N. ampullaria and N. bicalcarata do have one trait that separates them from the other Nepenthes species. They have fully paniculate (branched) inflorescences rather than the racemose-like (unbranched or almost unbranched) inflorescences more typical of the genus. Ancistrocladus, Triphyophyllum and related species also have paniculate inflorescences so we could assume the ancestral Nepenthes had paniculate inflorescences as well.
We know a little more about the ancestral Nepenthes species beyond what it would look like. All Nepenthes species tested are octoploids (8 copies of each chromosome) with 2n=80 (Heubl and Wistuba (1995), Lowrey (1991)) but cytologically appear to be diploid. Lowrey (1991) confirmed that gene copy level of commonly tested isozymes was diploid which means the alternate copies of the isozymes were lost or became inactive. It would take a more detailed study to determine the full level of gene diploidization.
There are many ways to form polyploids. The most common way in this group of plants is via hybridization between not quite compatible species that produce mostly sterile progeny. I say mostly sterile because with a small probability they produce aneuploid gametes that when the right combination unites, produce offspring of a higher, more stable, fertile ploidy level. Colchicine is used to induce this natural phenomenon at high frequency. The doubling has to happen three times: twice separately to get two different tetraploids then they have to cross and that hybrid doubles to get an octoploid. Evolutionarily these higher ploidy levels give the resulting plants a huge amount of genetic variation allowing them to develop new forms and functions. The high ploidy levels also present a physiological cost to the plant which is why over time most of the extra genes and quite often whole chromosomes are lost. Becoming polyploid is easy, dealing with or taking advantage of the consequences takes time. A long time. And since Drosophyllum is diploid the events had to happen after the Nepenthes line split from the Drosophyllum/Triphyophyllum line. Add to this that all Nepenthes species and hybrids appear to successfully interbreed and produce fertile offspring and you get the picture of recently evolved lineage with a relatively stable genome. Whatever happened to produce these details of the Nepenthes genome was essentially complete by the time the current radiation of species happened.
During the period between the split with its relatives and the radiation of the known Nepenthes species, something else genetic happened besides polyploidy. Nepenthes switched from having hermaphrodite flowers to being dioecious. That is its relatives have flowers containing both male and female parts. Nepenthes plants produce either male or female flowers but not both. And the flowers are very male or very female; there is no vestige left of the other sexual parts. This is not common among flowering plants. About 10% of flowering plant species are gender dimorphic and but less than 1% are are fully dioecious (See Barrett 2002 and Wikipedia Plant Sexuality for a review).
A little is known about how to evolutionarily get from from having both sex flowers to single sex flowers. Miller and Venable (2000, 2002) found an example of such a transition in the genus Lycium (Boxthorn, Solanaceae) facilitated by polyploidy. Most Lycium species are self-incompatible (the female part of the flower kills the pollen with the same genotype as its own to avoid self fertilization, see Wikipedia Self-incompatibility for more information) but some species are gender dimorphic; in this case having either female flowers or hermaphrodite flowers. Miller and Venable found the gender dimorphic species are polyploid and hypothesized that doubling the chromosomes disrupted the self-incompatibility system and created a situation were a male sterile mutation was able to spread throughout the population producing essentially female plants and both-sex plants.
This is an example of how our genetic system can cause seemingly bizarre things to happen evolutionarily (McDonald 1974). At the time of the self-incompatibility disruption there is a window of opportunity for sex-specific sterility genes to invade and/or persist in the population leading to sexual dimorphism. Until there has been time for the gene frequencies to reach an equilibrium, male sterile (i.e. female) plants would experience much less inbreeding depression and thus have higher fecundity and produce more healthier offspring than plants that are hermaphrodite and self pollinate. This would be the first step in the evolution of dioecious plants. The next step would be for the hermaphrodite flowers to become female sterile (i.e. male) and thus not put the energy into seeds that will not contribute much to the next generation. Of course the species will commit suicide if all the plants are male sterile or female sterile so the system needed to evolve with a proper balance and controls for there to be Nepenthes today.
Could the Nepenthes ancestor have been self-incompatible? Self-incompatibility is very common in flowering plants with some estimates on the order of 50% of species. Current dogma says evolution of self-incompatibility is very rare while the disruption of self-incompatibility is relatively common. Self-incompatibility continues to be common because self-incompatible species can maintain more genetic variation and are more likely to produce new species. Since many Drosera species in Australia are self-incompatible it is likely the ancestor of this whole group was self-incompatible. Self-incompatibility was lost in "simple" ways multiple times in Nepenthes relatives, something uncommon happened with Nepenthes.
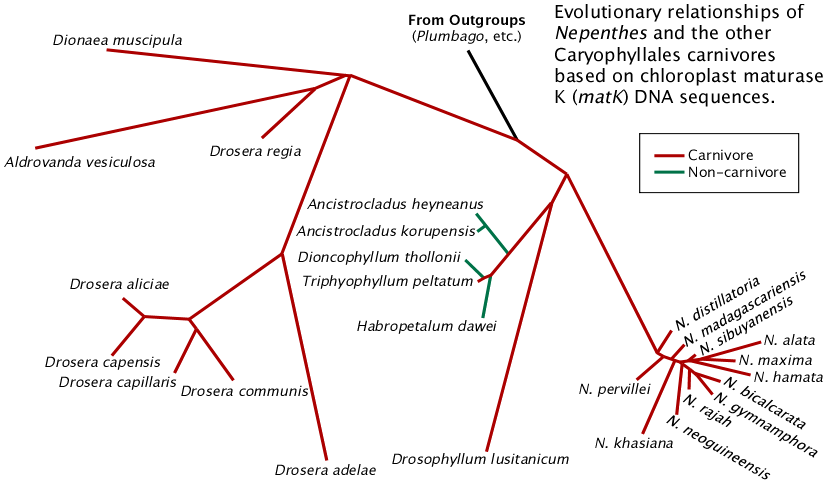
So a lot had to happen genetically to Nepenthes after its split with its ancestors. And we have not even gotten to the most obvious feature of the genus, its pitchers. (This will be covered separately.) All these evolutionary changes take time. Lots of time. How much time are we talking about? To put dates on the Nepenthes phylogeny we need fossils. For these plants the relevant fossils are mostly pollen. The Wikipedia Droseridites page has a good summary of the pollen data associated with Nepenthes. It is apparent that in the 35 to 65 million year old time frame it is difficult to link the pollen fossils to existing genera so we end up with, "well, it is most like Nepenthes but it could be Drosera". For all we know, the fossils could all be representatives of totally extinct lineages. What they do indicate is proto-Nepenthes-like plants were abundant along the northern Tethys Sea in what is now Europe and southwestern Asia. However there are candidates in other locations as well. An additional fossil of interest in this time frame is a winged fruit similar to Triphyophyllum named Dioncophyllites amurensis Fedotov. It was found in central Asia. Manchester, et al. (2009) document a number of east Asian endemic plants with fossil records along the northern Tethys. Similar studies should also be done for Africa.
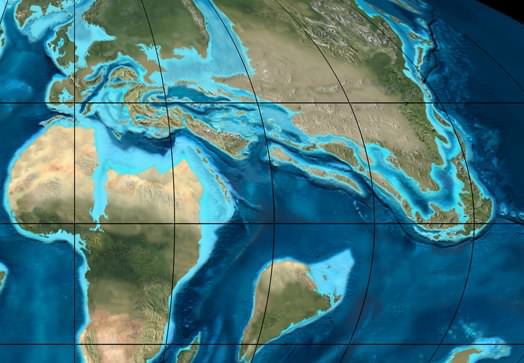
If we assume Nepenthes and the related genera Drosophyllum, Triphyophyllum, Ancistrocladus, Habropetalum and Dioncophyllum or their progenitors originated along the Tethys Sea and the locations of these plants today are the results of migrations from the Tethys, the pompom phylogenies make sense. We basically lost all the Tethys ancestors from climate change as the continents migrated and the Tethys Sea disappeared. This would mean Nepenthes as we know it evolved in what is now Europe and/or southwest Asia and at one point a fully modern Nepenthes arrived in the region of what is today the Indian Ocean. Similarly the progenitor or progenitors of Ancistrocladus and Triphyophyllum / Habropetalum / Dioncophyllum migrated to Africa. Nepenthes pompomed some time after arriving at its new home. Ancistrocladus did the same to become quite widespread across Africa and southeast Asia including areas where Nepenthes is found but not necessarily immediately after leaving Europe.
An alternative hypothesis, Nepenthes out-of-Africa, proposes that Nepenthes took the India taxi from southern Africa to Asia with the basal species getting off the taxi as Madagascar and the Seychelles split from India. For this to work fully modern Nepenthes would have had to exist over 100 million years ago. If they had existed at that time in that location we would expect to find Nepenthes in South America and Africa today along with other plants that were in that area at that time. Even if we assume they were there and are now lost or not found yet, a date this old is not concordant with the fossil dates or with the pompom configuration of the phylogenetic tree.
The out-of-Europe hypothesis makes the European Drosophyllum lusitanicum quite interesting. It would be very easy to confuse all the other species we have been discussing if you ignore the pitchers on Nepenthes and the occasional sundew-like leaves on Triphyophyllum. The plants are all more or less tropical lianas with similar leaves, stems, and flowers (if you squint). Without Triphyophyllum and its sundew-like leaves, the presence of the sundew-like Drosophyllum in this clade would stand out as being most bizarre. A sundew in the middle of clade of lianas? But unlike the surviving lianas, Drosophyllum did not migrate out of Europe. Europe is no longer a liana kind of place. Drosophyllum apparently adapted to the changed climate in Europe by losing all the no longer useful liana characters and taking the form of a full-on sundew.

Triphyophyllum peltatum with three glandular leaves in the greenhouse of the Botanical Gardens of Bonn. Cover photo from September 2010 CPN issue containing the article Rembold, Katja, Andreas Irmer, Simon Poppinga, Heiko Rischer and Gerhard Bringmann (2010) Propagation of Triphyophyllum peltatum (Dioncophyllaceae) and observations on its carnivory. Carniv. Pl. Newslett. 39(3):71-77. Photo by Katja Rembold.

Nepenthes inflorescences. Left: Fully paniculate inflorescence of Nepenthes pervillei (photo © Urs Zimmermann). Right: Racemose-like inflorescence of Nepenthes ventricosa. Fully paniculate flowers appear to be an ancestral feature of Nepenthes. Recent species have intermediate to racemose-like paniculate inflorescences. This may reflect a change in pollinators experienced by early migrants into the Borneo region.
In the taxonomy literature for the Caryophyllales carnivores there are references to cyme, panicle, raceme, and probably other inflorescence forms. Although these terms may have specific definitions that some taxonomists and plant anatomists might agree to (Wikipedia) the plants could not care less about definitions and do whatever works for them. It is not clear Nepenthes inflorescences fit any of the strict definitions exactly but panicle is closest. Raceme appears to be used most for inflorescences typical for Drosera where the flowers open in a wave going up the stem—never mind Nepenthes does that too but it is more subtle. Evolutionarily, changes in inflorescence structure and the pattern of flower opening generally involve simple changes in gene regulatory sites coordinating when, where, and how much hormone genes are expressed.

Nepenthes maxima female flowers. Photo by Mach Fukada.

Nepenthes flowers. Left: Female flower of Nepenthes pervillei. Photo © Urs Zimmermann. Right: Male flower of Nepenthes ventricosa. All Nepenthes plants are either female or male—in botanical terms they are dioecious. There is variation in the configuration of the flowers between species but there is no vestige of the opposite sex in any species. The evolution of dioecious flowers must have happened before any of the currently known species came into existence.

Nepenthes pervillei on the Seychelles Islands between India and Africa. Photo © Urs Zimmermann.
The Tethys Sea is between Africa and the Europe/Asia continent in this 65 million year old "satellite view". One possible location for the evolution Nepenthes is the northern part of the Tethys Sea in what is now Europe. Image © Ron Blakey, Northern Arizona University Geology.
As Africa collided with Europe, many species that evolved on the islands in the Tethys Sea escaped to Africa and Asia. Many that stayed home are no longer with us. This view is from the Oligocene, 35 million years ago. Image © Ron Blakey, Northern Arizona University Geology.

Drosophyllum lusitanicum. Photo by Tom Cahill.
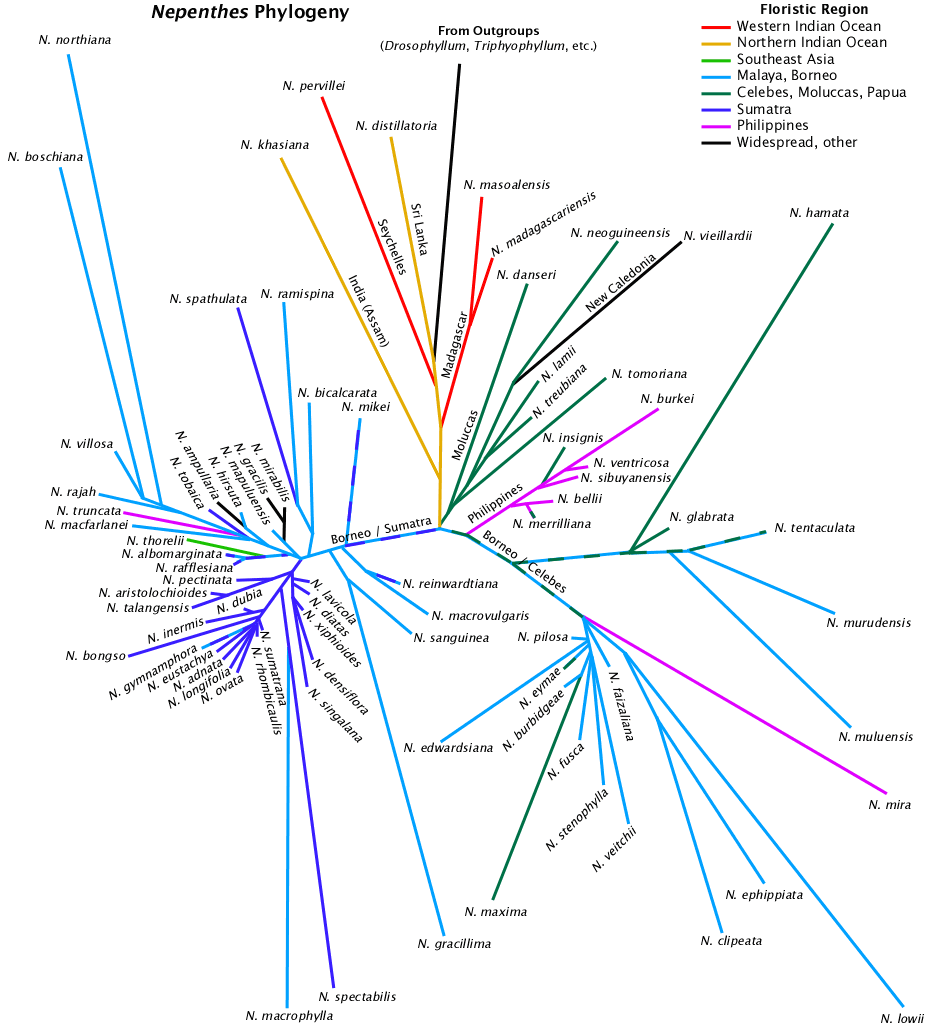
The data used for this tree are chloroplast trnK gene intron, matK sequences from Heubl, Meimberg, and associates (see references) greater than 2000 bases long and deposited with NCBI. Care was taken to exclude the translocated matK pseudogene sequences in the figure. Analysis was via NCBI Blast, graphed via PHYLIP drawtree unrooted tree drawing program and detailed manually in a drawing program. The lengths of the lines are an indication of genetic distance. Triphyophyllum peltatum, Drosophyllum lusitanicum, and a number of Ancistrocladus species were used as the outgroup.
When two samples of the same species were included in the analysis, the one with the best quality sequence was selected for the figure.
Some samples may be misidentified as to species. "N. alata" was not included because the source location information given in Meimberg et al. (2001) was impossible for that species. There were no clarifications or corrections in later publications where the same data were used. Although the probability is small that "N. thorelii" is really N. thorelii, the plant used for the study could be a related species although it could be a hybrid outside its species group. It is left it in for the sake of discussion.
Specific details of the phylogeny could change with additional species or analysis with a different program. The data used for the tree are raw sequence data not cleaned to removed artifacts.
The tree contains four species marked as "not accepted" in the Carnivorous Plant Names Database:
| CP Names Database reason for "not accepted" | Phylogeny support for CP Names Database assertion |
| N. ramispina = N. gracillima | not supported, the sequences are different |
| N. pectinata = N. gymnamphora x N. singalana | not supported, the sequences of all three are different |
| N. pectinata = N. ovata | not supported, the sequences are different |
| N. xiphioides = N. gymnamphora | not supported, the sequences are different |
| N. murudensis = N. tentaculata x N. reinwardtiana | equivocal, sequence close to N. tentaculata but not identical, different from N. reinwardtiana. A more detailed study with more sequences from multiple individuals might confirm hybrid origin. |
In four cases direct examination of the sequence data do not support the assertion in the database. In one it is too close to call. The classification problems could be inadequate description of the species.
Floristic region data used are from the Carnivorous Plant Names Database and are grouped by geographic association. Floristic regions may not be fully concordant with geographic features or locations of the same or similar names.
The dioecious nature of Nepenthes puts some fairly strong constraints on the migration of species to new locations. One seed of a plant with flowers that can self pollinate is enough to found a whole new branch of the species at some distant location. But one seed of Nepenthes is not enough. The plant will be either male or female. If it is male there is no hope of producing seeds on its own. If female it might produce seeds without pollination but they should be clones and thus females as well. Since Nepenthes plants are potentially immortal they can hang out for a long time and maybe a second seed of the opposite sex will happen along. Maybe very very rarely a plant will produce flowers of both sexes. It is of course possible two or more seeds will hitchhike to the same place at the same time.
On the other hand since all Nepenthes species can interbreed and in fact function genetically like one hyper-variable species, one seed can bring in a lot of new variation if it establishes a plant among a population of another species. This can lead to the establishment of new taxonomic characters in that population leading humans to name that population as a new species. This has undoubtedly happened many times in Nepenthes so a phylogenetic tree produced from nuclear genes would probably lay out differently from the tree above which is based on chloroplast genes. Meimberg et al. (2006) did a limited study of nuclear genes and did in fact find some species that showed different associations. I do not feel confident enough in the public data to reproduce it here.
The chloroplast phylogenetic tree for Nepenthes is consistent with an initial population of Nepenthes in India and/or southwest Asia that gave rise to the species now in Madagascar and the Seychelles via long distance dispersal. The species now in Sri Lanka and Assam could be more direct remnants of that ancestor with the Sri Lanka population isolated very early. It would be incorrect to say any of the exiting species today is the ancestor although with the current analysis N. khasiana is the last species to branch off before the ancestor infested the Malay Peninsula and Borneo founding the bulk of the genus.
It appears there were two or more introductions into the Malay/Borneo region or if it was one introduction there was a split into at least four branches soon after. One branch was probably on the west side of Borneo and the others on the coast and in the islands on east side of Borneo. If this event happened 20 to 35 million years ago simple within-landmass dispersal would be sufficient to stage the species near where their future homes would appear. If it happened later, longer distance dispersal over a few hundred km of ocean could have established major branches of the eastern population in Sulawesi, the Maluku (Moluccan) Islands, and the Philippines when they were closer to Borneo than they are today. So how could all this movement happen?
Nepenthes seeds are typically wind dispersed. If you have ever handled the seeds or tried to make seed packets you will know they get everywhere. Typical strong storm winds will probably move seeds on the order of a km. However it is quite reasonable that major tropical storms will disperse seeds over distances on the order of hundreds of km. Can this account for what we can deduce from the phylogeny? Yes it does for clusters of species on large landmasses like the initial migration around proto-Borneo and the more recent invasion of high elevations. But it does not work for the very long distances seen in the phylogeny.
During periods of low sea level such as during the recent Pleistocene ice ages it is quite possible Nepenthes species spread across the land bridges. This may account for the distribution of a few species but it is not a general feature of Nepenthes range expansion and evolution. N. albomarginata, N. rafflesiana, N. eymae, N. tentaculata, and N. gymnamphora might fit this scenario but they may have already been in place before the land bridges.
The most likely agent for long distance dispersal of Nepenthes is migrating shorebirds and seabirds. During their migration the birds will stop in coastal marshes to recharge before moving on. Some islands also have large rookeries of seabirds. It is hard to explain the presence of Nepenthes on Madagascar, the Seychelles, New Guinea, New Caledonia, and other islands such as Palau in Micronesia any other way. What is most astonishing is the range over which N. mirabilis is found. Most Nepenthes species have very small ranges. A range map for N. mirabilis would include almost everywhere other Nepenthes are found in south east Asia and then include an area well beyond that. Charles Clark reports that even though this species is widespread it tends to live in coastal swamps where few if any other Nepenthes species are found. It would be interesting to know if N. mirabilis has special adaptations to facilitate migration or if just being in coastal swamps is enough.
The dioecious nature of Nepenthes limits migration. Does having separate sexes also explain the large number of species in a relatively small area? It does. The impossibility of self pollination in Nepenthes allows small populations to maintain large amounts of genetic variation relative to species that can inbreed. With this variation the populations can develop characters that taxonomists would use to define species. So if the explosion of species in Sumatra of the clade with N. lavicola at the base resulted from the rise of the Barisan Mountains, both isolation between populations on new mountains and the generation of new habitats gave great opportunity for Nepenthes to create new forms. A selfing species would be less likely to be able to maintain that much variation in characters. That is not to say it could not happen. With selfing species we might refer to the variant populations as "ecotypes". With Nepenthes each ecotype or population could have enough distinguishing taxonomic characters to be referred to as separate species. This results in each mountain having its own species of Nepenthes while a patchy but widespread self-compatible Drosera species such as D. ultramafica growing in association with Nepenthes on mountain tops shows little variation between distant locations.
-- John Brittnacher
May 2011
For a more detailed discussion please see the following articles and articles they reference.
Heubl, G. and Wistuba, A. (1995) A cytological study of the genus Nepenthes L. (Nepenthaceae). Sendtnera 4:169-174. (Note the article says a ploidy level of "16x (8x?)". To get 16x they were making the assumption that the basal chromosome level for the Caryophyllales carnivores is x=5. It is quite possible it is 5 or 6.)
Lowrey, T. K. (1991) Chromosome and isozyme number in the Nepenthaceae. Am. J. Botany 78(6) Suppl.: 200-201.
Barrett, S. C. H. (2002) Evolution of sex: The evolution of plant sexual diversity. Nature Reviews Genetics 3:274-284.
Miller, J. S. and Venable, D. L. (2000) Polyploidy and the evolution of gender dimorphism. Science 289:2335–2338.
Miller, J. S. and Venable, D. L. . (2002) The transition to gender dimorphism on an evolutionary background of self-incompatibility: an example from Lycium (Solanaceae). Am. J. Botany 89(2):1907-1915.
McDonald, J. F. (1974) Can Species Commit Suicide? Journal of Paleontology 48(3):565-567.
Meimberg, H., Wistuba, A., Dittrich, P., and Heubl, G. (2001) Molecular phylogeny of Nepenthaceae based on cladistic analysis of plastid trnK intron sequence data. Plant Biology 3:154-175.
Heubl, G., G. Bringmann, and H. Meimberg (2006) Molecular phylogeny and character evolution of carnivorous plant families in Caryophyllales – revisited. Plant Biol. 8:821-830.
Meimberg, H. and Heubl, G. (2006) Introduction of a Nuclear Marker for Phylogenetic Analysis of Nepenthaceae. (2006) Plant Biology 8:831–840
Meimberg, H., Thalhammer, S., Brachmann, A., and Heubl, G. (2006) Comparative analysis of a translocated copy of the trnK intron in the carnivorous genus Nepenthes (Nepenthaceae). Molecular Phylogenetics and Evolution 39:478–490.
Manchester, S. R., Chen, Z.-D., Lu, A.-M. and Eumura, K. (2009) Eastern Asian endemic seed plant genera and their paleogeographic history throughout the Northern Hemisphere. Journal of Systematics and Evolution 47:1-42.
Clarke, C. M. (1997) Nepenthes of Borneo. Natural History Publications (Borneo), Kota Kinabalu.
Clarke, C. M. (2001) Nepenthes of Sumatra and Peninsular Malaysia. Natural History Publications (Borneo), Kota Kinabalu.
McPherson, S.; Edited by A. Fleischmann and A. Robinson. (2010) Carnivorous Plants and Their Habitats: Volume 1 and Volume 2. Redfern Natural History Productions Ltd., Poole, GB

Nepenthes alba growing on the bleak, windswept summit of Mount Tahan on the Malay Peninsula. Photo by Stewart McPherson (2009) Carniv. Pl. Newslett. 38(4) p. 105.
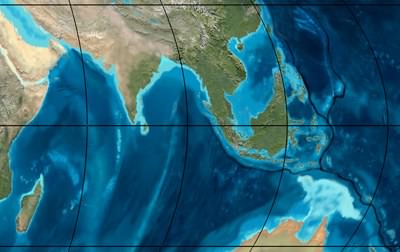
South east Asia during the Miocene, about 20 million years ago. New Guinea did not exist. Sumatra and the Malay Peninsula were not differentiated and were connected to Borneo. Borneo was also connected to Sulawesi and possibly to the Philippines. Image © Ron Blakey, Northern Arizona University Geology.
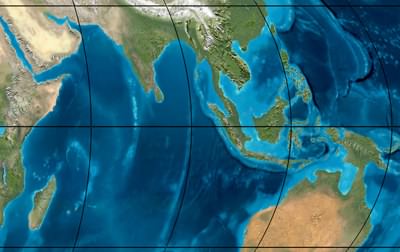
South east Asia during the Pleistocene, about 50 thousand years ago. The image shows a land bridge from Sumatra to the Malay peninsula and from New Guinea to Australia. Image © Ron Blakey, Northern Arizona University Geology.

Nepenthes sibuyanensis from the Philippines. Photo by Phill Mann (1998) Carniv. Pl. Newslett. 27(1) front cover.